Our
TRPV (Vanilloid); TRPM; TRPA and TRPCs have proven excellent for studying
Transient receptor potential ankyrin 1 (TRPA1) is activated by noxious cold (<17°C) and contributes to cold and mechanical hypersensitivity after inflammation and nerve injury:
Yun Sook Kim, PhD, Hoon Kap Jung, DDS, Tae Kyung Kwon, DDS, Chin Soo Kim, DDS, PhD, Jin Hyun Cho, DDS, PhD, Dong Kuk Ahn, DDS, PhD, Yong Chul Bae, DDS, PhD.Expression of Transient Receptor Potential Ankyrin in Human Dental Pulp. Journal of Endodontics. Available online 8 June 2012. doi.org/10.1016/j.joen.2012.04.024.
Highlights: TRPA1 was expressed in a large number of axons branching extensively in the peripheral pulp and in a few axons within the nerve bundles in the core of the coronal pulp and in the radicular pulp. Under electron microscopy, TRPA1 immunoreactivity was typically localized near the plasma membrane of unmyelinated axons in the peripheral pulp, suggesting that in these axons it may act as a functional receptor. The proportion of axons expressing TRPA1 in neurofilament 200–positive axons significantly increased in the painful pulp compared with the normal pulp. TRPA1 was also densely expressed in the processes and the cell body of odontoblasts. A large number of axons coexpressed TRPA1 and Nav1.8.
Images: Immunofluorescent staining for (A) TRPA1 in the human normal dental pulp is completely abolished by (B) preadsorption with a control peptide, proving the specificity of the TRPA1 antiserum (×200, scale bars = 50 µm). doi.org/10.1016/j.joen.2012.04.024.
Related Reagents:
All TRP Antibodies
Pain and Inflammation Research Antibodies
Neurotransmission -Neurotransmission Research Antibody Categories
I have had to say to many customers, "our guinea pig P2x3 is on backorder". The increasing number of pubs referencing this antibody only amped demand.
We tried and tried to re-make it. The result was none of the bleeds we tested had a signal strong enough to release the antibody. We had a customer suggest re-testing several of the more promising bleeds. Thank you! We have good news on results and we are offering for 50% off. This is to acknowledge the investment required for TSA and Guinea Pig Biotinylated Antibody.
![]() Here're the recent pubs I referenced:
Gabriela Castañeda-Corral, Héctor I. Rocha-González, Beatriz Godínez-Chaparro, Juan Miguel Jiménez-Andrade and Vinicio Granados-Soto. Role of the spinal Na+/H+ exchanger in formalin-induced nociception. Neuroscience Letters. doi:10.1016/j.neulet.2011.06.048.... SP (guinea pig; 1:500; Cat# GP14110; Neuromics), CGRP (goat, 1:500; Cat# Ab36001; Abcam) and P2X3 receptor (guinea pig: 1:10,000; Cat# GP10108; Neuromics)...
Anna M.W. Taylora and Alfredo Ribeiro-da-Silva. GDNF levels in the lower lip skin in a rat model of trigeminal neuropathic pain: Implications for nonpeptidergic fiber reinnervation and parasympathetic sprouting. PAIN Volume 152, Issue 7, July 2011, Pages 1502-1510. doi:10.1016/j.pain.2011.02.035.
...Sections were then incubated for 48h at 4°C with a guinea pig polyclonal anti-P2X3 (1:25,000; Neuromics, Edina, MN)...
Bone crushing pain. This describes pain of the highest order. Our friend, Dr. Joseph Ghilardi, VAMC-Mpls. and his colleague, Dr. Patrick Manthy are finding the root causes of the intense and growing pain suffered by Cancer Victims. Here are highlights of a recent study:
Pain frequently accompanies cancer. What remains unclear is why this pain frequently becomes more severe and difficult to control with disease progression. Here we test the hypothesis that with disease progression, sensory nerve fibers that innervate the tumor-bearing tissue undergo a pathological sprouting and reorganization, which in other nonmalignant pathologies has been shown to generate and maintain chronic pain. Injection of canine prostate cancer cells into mouse bone induces a remarkable sprouting of calcitonin gene-related peptide (CGRP+) and neurofilament 200 kDa (NF200+) sensory nerve fibers. Nearly all sensory nerve fibers that undergo sprouting also coexpress tropomyosin receptor kinase A (TrkA+). This ectopic sprouting occurs in sensory nerve fibers that are in close proximity to colonies of prostate cancer cells, tumor-associated stromal cells and newly formed woven bone, which together form sclerotic lesions that closely mirror the osteoblastic bone lesions induced by metastatic prostate tumors in humans. Preventive treatment with an antibody that sequesters nerve growth factor (NGF), administered when the pain and bone remodeling were first observed, blocks this ectopic sprouting and attenuates cancer pain. Interestingly, reverse transcription PCR analysis indicated that the prostate cancer cells themselves do not express detectable levels of mRNA coding for NGF. This suggests that the tumor-associated stromal cells express and release NGF, which drives the pathological reorganization of nearby TrkA+ sensory nerve fibers. Therapies that prevent this reorganization of sensory nerve fibers may provide insight into the evolving mechanisms that drive cancer pain and lead to more effective control of this chronic pain state.  . J. Neurosci., Nov 2010; 30: 14649 - 14656 ; doi:10.1523/JNEUROSCI.3300-10.2010
Here're several other pubs referencing use of our antibodies in studying bone cancer pain:
Kyle G. Halvorson, BA, Molly A. Sevcik, BA, Joseph R. Ghilardi, BS, BA, Lucy J. Sullivan, BA, Nathan J. Koewler, BS, Frieder Bauss, PhD, and Patrick W. Mantyh, PhD. Intravenous Ibandronate Rapidly Reduces Pain, Neurochemical Indices of Central Sensitization, Tumor Burden, and Skeletal Destruction in a Mouse Model of Bone Cancer. Published online 2008 April 14. doi: 10.1016/j.jpainsymman.2007.10.005
...pro-dynorphin (DYN, polyclonal guinea pig anti-rat, 1:1,000; Neuromics, Minneapolis, MN)...
Timothy K. Y. Kaan, Ping K. Yip, Sital Patel, Meirion Davies, Fabien Marchand, Debra A. Cockayne, Philip A. Nunn, Anthony H. Dickenson, Anthony P. D. W. Ford, Yu Zhong, Marzia Malcangio, and Stephen B. McMahon Systemic blockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour in rats. Brain, September 2010; 133: 2549 - 2564.
......Slides were then incubated with rabbit anti-P2X3 (1:2000, Neuromics) and sheep anti-calcitonin gene-related peptide (1:1000, Biomol...anti-beta-III-tubulin (1:4000, Promega) and guinea pig anti-P2X3 (1:100, Neuromics). The next day, after three washes with phosphate-buffered......
I will keep you posted on this important topic.
Our Opioid Receptor Antibodies have set a potent standard for studying Nociceptive and Neuropathic Pain. Related Publications.
We want to recognize Dr. Zaijie Jim Wang and his team for being the first to use our Mu Opioid Receptor for studying the potential role of microRNAs in Nociception.
Ying He, Cheng Yang, Chelsea M. Kirkmire, and Zaijie Jim Wang. Regulation of Opioid Tolerance by let-7 Family MicroRNA Targeting the µ Opioid Receptor. The Journal of Neuroscience, July 28, 2010, 30(30):10251-10258; doi:10.1523/JNEUROSCI.2419-10.2010
Abstract: MicroRNA has emerged as a critical regulator of neuronal functions. This study aimed to test whether let-7 microRNAs can regulate the µ opioid receptor (MOR) and opioid tolerance. Employing bioinformatics, we identified a let-7 binding site in the 3'-untranslated region (UTR) of MOR mRNA, which was experimentally confirmed as a direct target of let-7. The repressive regulation of MOR by let-7 was revealed using a LNA-let-7 inhibitor to knockdown let-7 in SH-SY5Y cells. Conversely, morphine significantly upregulated let-7 expression in SH-SY5Y cells and in a mouse model of opioid tolerance. The LNA-let-7 inhibitor decreased brain let-7 levels and partially attenuated opioid antinociceptive tolerance in mice. Although chronic morphine treatment did not change overall MOR transcript, polysome-associated mRNA declined in a let-7-dependent manner. let-7 was identified as a mediator translocating and sequestering MOR mRNA to P-bodies, leading to translation repression. These results suggest that let-7 plays an integral role in opioid tolerance.
- Western blot analysis. Western blot analysis was performed as previously described (Tang et al., 2006) using the anti-µ opioid receptor antibody (1:1000; Neuromics). The expression of β-actin was similarly determined from the same blots using a monoclonal antibody (1:10,000; Sigma).
- For immunofluorescence analysis, the antibody for hDcp1a (Santa Cruz Biotechnology) and MOR were used at 1:500 and 1:5000 dilutions, respectively. Secondary anti-goat and anti-mouse antibodies labeled with Alexa 488 and Alexa 594 fluorochromes (Invitrogen), respectively, were used at 1:500 dilutions.
Dr. Tomas Hokfelt and his team at Karolinska Institute recently published use of our Opioid Receptor Antibodies and Substance P Antibody.They show the interplay of DOR and MOR in modulation of nociceptive afferent transmission and opioid analgesia.
Hai-Bo Wanga, Bo Zhaoa, Yan-Qing Zhonga, Kai-Cheng Li, Zi-Yan Li, Qiong Wang, Yin-Jing Lua, Zhen-Ning Zhang, Shao-Qiu He, Han-Cheng Zheng, Sheng-Xi Wu, Tomas G. M. Hökfelt, Lan Baob, and Xu Zhanga. Coexpression of δ- and μ-opioid receptors in nociceptive sensory neurons. PNAS July 20, 2010 vol. 107 no. 29 13117-13122.
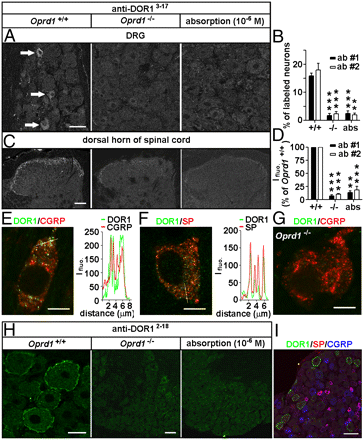
Immunostaining. Adult rats, mice, and Oprd1 exon 1-deleted mice were fixed. Cryostat sections of L4 and L5 DRGs and spinal cord segments were processed for immunofluorescence staining (13) with Rb anti- DOR13–17 (1:2,000–1:60,000; DiaSorin and 1:4,000–1:60,000; Neuromics), Rb anti-DOR12–18 (1:30,000–1:120,000; Alomone), Rb anti-DOR1358–372 (1:1,000–1:2,000; Lifespan Biosciences), Rb anti- MOR (1:1,000; Neuromics); guinea pig anti- SP (1:500; Neuromics), and mouse anti-CGRP (1:1,000; Biogenesis) antibodies. IB4-labeling was carried out with fluorescein-labeled GSL I-IB4 (1:200). The Myc-DOR1–transfected HEK293 cells and neurons were fixed and processed with mouse anti-Myc antibodies (1:500; DSHB). Nuclear DAPI staining was used to indicate HEK293 cells in control experiments.

Images: Distinct distribution patterns of DORs in subsets of DRG neurons of mice. Immunostaining with antibodies against DOR13–17 [A: 1:30,000, antibody 1 (ab #1); DiaSorin and C: antibody 2 (ab #2); Neuromics] shows DORs in small DRG neurons and afferent fibers in spinal laminae I–II. This immunostaining pattern is abolished by the antiserum preabsorption or the deletion of Oprd1 exon 1. Reduction in immunostaining is quantitatively assayed by determining the percentage of positive DRG neurons (B; n = 6) and fluorescence intensity (Ifluo.) in the laminae I–II (D; n = 5). **P < 0.01; ***P < 0.001. (Scale bars: A and C, 40 μm.). DOR labeling (anti-DOR13–17, 1:30,000; DiaSorin) associated with vesicles in peptidergic small DRG neurons (E and F) is absent in Oprd1 exon 1-deleted mice (G). Colocalization of DORs and neuropeptides is shown by correlated peaks of Ifluo. measured along lines. (Scale bar: 8 μm.) (H) Immunostaining with antibodies against DOR12–18 (1:60,000; Alomone) shows the presence of DORs on the cell surface of large DRG neurons of mice. (Scale bar: 25 μm.) This staining pattern is abolished by preabsorption and is absent in Oprd1 exon 1-deleted mice. (Scale bar: 80 μm.) (I) Triple-immunostaining shows that DOR+ large DRG neurons contain neither SP nor CGRP. (Scale bar: 80 μm.)
Immunoblotting.The samples were processed for SDS/PAGE, transferred, probed with Rb antibodies against MOR (1:500; Neuromics), phospho-DOR1 (1:1,000; Neuromics), phospho-MOR (1:1,000; Neuromics), Myc (1:500; DSHB), Flag (1:1,000; Sigma), or actin (1:50,000; Chemicon) and visualized with enhanced chemiluminescence (19).
Neuromics' Pain Research Customers continue to make gains using our Pain and Inflammation Research Antibodies and Transfection Kits. Here are the latest pubs: Hua Zhang and A. S. Verkman. Aquaporin-1 Tunes Pain Perception by Interaction with Nav1.8 Na+ Channels in Dorsal Root Ganglion Neurons. February 19, 2010 The Journal of Biological Chemistry, 285, 5896-5906.
...chicken anti-calcitonin gene-related peptide (CGRP; 1:500, Neuromics, Edina, MN)... Nathaniel A. Sowa, Bonnie Taylor-Blake, and Mark J. Zylka. Ecto-5'-Nucleotidase (CD73)  Inhibits Nociception by Hydrolyzing AMP to Adenosine in Nociceptive Circuits. The Journal of Neuroscience, February 10, 2010, 30(6):2235-2244; doi:10.1523/JNEUROSCI.5324-09.2010. Inhibits Nociception by Hydrolyzing AMP to Adenosine in Nociceptive Circuits. The Journal of Neuroscience, February 10, 2010, 30(6):2235-2244; doi:10.1523/JNEUROSCI.5324-09.2010. ...rabbit anti-P2X3-RA10109, Neuromics; 1:750), rabbit anti- VR1 C-Terminus (TRPV1) - mouse specific (RA14113, Neuromics; 1:750 Images: Images: Confocal images showing the effect of RTX on mu opioid receptor and TRPV1 immunoreactive DRG neurons and afferent terminals in the spinal cord. A: representative confocal images showing mu opioid receptor (green) and TRPV1 (red) immunoreactivities in DRG neurons of one vehicle- and one RTX-treated rat. Scale bar, 40 um. B: confocal images showing mu opioid receptor (green) and TRPV1 (red) immunoreactivities in afferent terminals in the spinal dorsal horn of 1 vehicle- and 1 RTX-treated rat. Scale bar, 80 um. Inset: high-magnification images (scale bar = 5 um) showing co-localization of mu opioid receptor and TRPV1 immunoreactivity in the lamina I. Co-localization of the mu opioid receptor and TRPV1 immunoreactivity is indicated in yellow when 2 images are digitally merged. All images are single confocal optical sections. Shao-Rui Chen and Hui-Lin Pan. Loss of TRPV1-Expressing Sensory Neurons Reduces Spinal mu-Opioid Receptors But Paradoxically Potentiates Opioid Analgesia. doi:10.1152/jn.01343.2005.
Neuromics' i-Fect ™ siRNA Transfection Reagent continues to be used a tool for studying expression of genes suspected to paly a role in pain . Expression studies include: DOR , The β3 subunit of the Na+,K+-ATPase, rSNSR1, NTS1. NAV1.8, CaV1.2 and more. Here's a link to all transfection publications: Transfection Kit Pubs
Related Reagents: |
|
Dr. Irmgard Tegeder and team have been using our VR1 N-Terminus (TRPV1) as a marker to study how B kinase (IKK) plays a role in sensory neuron excitability and nociception. The antibody was used for both Western Blot and Immunohistochemistry. Here's a recent publication of their results: Vanessa Bockhart, Cristina Elena Constantin, Annett Häussler, Nina Wijnvoord, Maike Kanngiesser, Thekla Myrczek, Geethanjali Pickert,Laura Popp, Jürgen-Markus Sobotzik, Manolis Pasparakis, Rohini Kuner, Gerd Geisslinger, Christian Schultz, Michaela Kress, and Irmgard Tegeder. Inhibitor B Kinase β Deficiency in Primary Nociceptive Neurons Increases TRP Channel Sensitivity. The Journal of Neuroscience, October 14, 2009, 29(41):12919-12929; doi:10.1523/JNEUROSCI.1496-09.2009 Related Reagents: All TRPV (Vanilloid); TRPM; TRPA and TRPC AntibodiesPain and Inflammation Research Antibodies
We would like to Dr. Andrew Todd for sharing this excellent IHC image using our Guinea Pig ProDynorphin (rat) antibody.  Image: Staining of adult rat spinal cord. The tissue is perfusion-fixed (4% freshly prepared formaldehyde) adult rat spinal cord, reacted overnight with the PPD at 1:1000 and then o/n in Alexa488 secondary (raised in donkey, Invitrogen, 1:500). The confocal image stack was taken through a 60x oil lens (Bio-Rad Radiance confocal) - pixel size is 0.196 micrometre and this is a projection of 10 confocal optical sections at 0.5 micrometre z-spacing. Customer PublicationsRelated Reagents: proDynorphin (guinea pig)Opioid ReceptorsPain and Inflammation Antibodies
|